Host-Parasitoid Spatial/Landscape Ecology
Behaviorally Based Landscape-Level Modeling
Dr. John Reeve and I have been collaborating on the development of behavior-based landscape-level models to address one of the most significant questions in ecology today – how does spatial heterogeneity affect the long-term temporal population dynamics and persistence of interacting species? The main objective is to develop a spatially realistic model that will reveal the mechanisms underlying the effects of habitat fragmentation and loss, and the invasion of exotic plant species, on the dynamics and persistence of a predator and its prey.
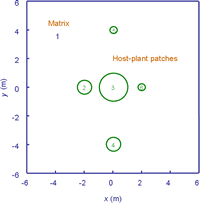
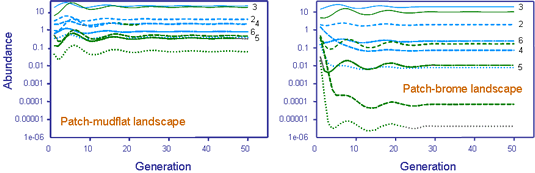
We have been developing such models for the planthopper (Prokelisia crocea) and its egg parasitoid (Anagrus columbi) residing in a landscape consisting of host-plant patches embedded in a heterogeneous matrix (Cronin & Reeve 2005, Reeve et al. 2008, Reeve and Cronin 2010, Xiao et al. 2013, Cronin and Reeve 2014), in which movement rates and boundary behavior vary with the composition of the matrix (Haynes 2004, Haynes & Cronin 2006). We briefly illustrate this approach on a hypothetical landscape using the software package FEMLAB 3.1 (COMSOL AB 2005).
The solution process consists of drawing the landscape and then specifying diffusion, oviposition, mortality, and parasitoid attack rates within each domain, as well as boundary behavior on the patch-matrix edge. The video below shows the movement of planthoppers across the landscape. Numerical solutions were obtained for two matrix types, mudflat and brome (a grass), using parameter values estimated from observations of P. crocea movement (Reeve et al. 2008).Similar models have been developed for A. columbi, based on a mark-recapture experiment (Reeve and Cronin 2010). The solutions illustrate the importance of matrix type, patch size, and edge behavior on host and parasitoid abundance. Densities were consistently higher for a cordgrass-mudflat (left) vs. cordgrass-brome landscape (right), because boundary behavior on the cordgrass-mudflat landscape retains dispersing insects within cordgrass patches (Cronin and Reeve 2014). Large cordgrass patches also had higher densities than small ones, as would be expected, and patches closer to the large central patch also had higher densities.