Host-Parasitoid Spatial/Landscape Ecology
Mainland-Island Metapopulation Structure

Through field experiments and a 7-generation census of numerous cordgrass patches, we were able to quantify the movement behavior of P. crocea and A. columbi and assess their spatial population structure (Cronin 2003a,b,c, 2004, Cronin et al. 2004). As was found with many insect species, planthopper within-patch movement was well described by a diffusion process. Emigration by planthoppers was also typical – emigration rate increased as the density of conspecifics or spiders increased, and decreased as patch size increased. Immigration by planthoppers was an increasing function of patch size, but was independent of patch isolation. The natural distribution of planthoppers in a prairie fragment was well predicted from the dispersal experiments. Planthopper densities and patch occupancy rates were positively correlated, and extinction rates negatively correlated with patch size, but patch isolation had no effect on any of these variables. Overall, local planthopper populations went extinct at a rate of 23% per generation; however, extinctions never took place in patches > 1 ha. Finally, the dynamics of local patch populations were asynchronous, even for patch pairs < 10 m apart.
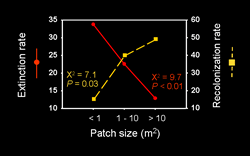

Because of the tiny size of A. columbi (0.7 mm), information on its movement has been challenging to obtaint. However, we have perfected a mark-recapture technique for this species (Cronin & Haynes 2004), and applied it to A. columbi (Reeve and Cronin 2010). Based on these data, it is clear that the movement behavior of A. columbi differs in important respects from that of its host (Cronin 2003a,c, 2004). Marked wasps strongly bias their movements towards the edge when released in matrix, and to a lesser extent when released in cordgrass, while edge releases showed little bias. Our results suggest the wasps are attracted to cordgrass patches across short distances, and that bias in their movements may concentrate them at the patch edge. Their edge behaviour is qualitatively different from that of the host insect. A. columbi emigration rate is neither dependent on host density nor its own density. However, parasitoids do suffer significant density dependent reductions in reproduction when they remain in patches bearing high conspecific densities. Perhaps the most important difference between the 2 species is that A. columbi is much less vagile than P. crocea. Immigration rates of A. columbi decrease exponentially with increasing patch isolation. An increase in plant density, decrease in host density and decrease in parasitoid density (lagged 1 generation) all increased the likelihood of A. columbi extinction within a patch (51% mean extinction rate per patch per generation). The dependency on multiple trophic levels may explain the higher extinction risk for the parasitoid than its host (Cronin 2004). As with the host, fluctuations in local population densities were asynchronous and populations of A. columbi in large patches never went extinct during the study. We have concluded that P. crocea and A. columbi exhibit mainland-island metapopulation dynamics among host-plant patches within a prairie landscape.





