Evolution and Ecology of Invasive Plant Species
Invasive Plant Species Affect Native Faunal Species Via Their Impact on the Connectivity Among Native Habitat Patches

The invasion and spread of exotic plant species is a major threat to population persistence and biodiversity in general. A plausible way in which exotic plant species might affect native faunas is via their effects on connectivity among native habitat patches (Cronin & Haynes 2004, Cronin 2007, Bezemer et al. 2014). Research by Kyle Haynes and me illustrates this point.
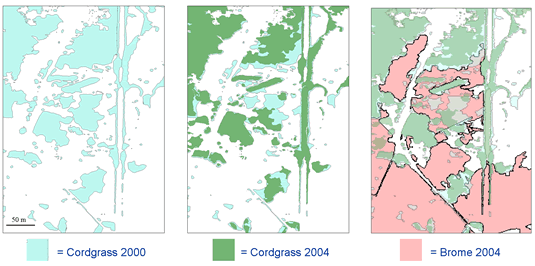
Prairie cordgrass (

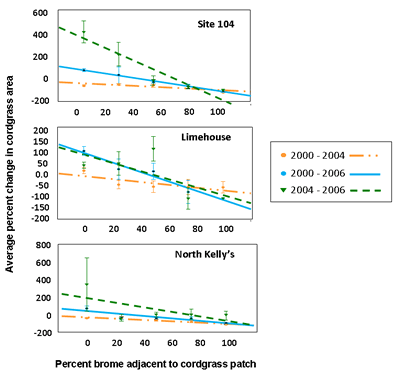
My former graduate student, Forrest Dillemuth and I used GPS and GIS technology to map the spread of brome and change in cordgrass patch distribution and abundance in three prairie fragments from 2000-2006 (Dillemuth et al. 2009). Over the course of 6 years, the structure of the landscape changed dramatically (see representative maps below ). Results from our GIS spatial analyses indicated that cordgrass patch growth was two times greater in areas not invaded by smooth brome versus areas that were heavily infested with smooth brome. In particular, cordgrass patches declined in size by an average of 61% in each prairie fragment. The change in cordgrass patch size was strongly correlated with the percentage of brome surrounding the patch – patches with a high percentage of neighboring brome had the greatest reduction in size (see Figure 1).
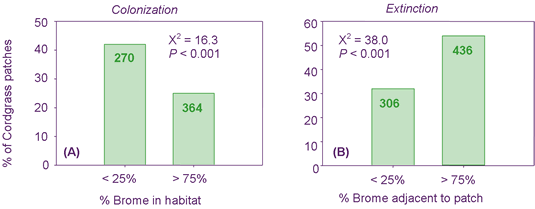
As shown in Figure 2, the probability of establishment of a new patch of cordgrass averaged 1.3 times higher in areas of low brome coverage (< 25%) than areas of high brome coverage (> 75%). Conversely, existing cordgrass patches were 7.8 times more likely to go extinct in areas of high than low brome coverage.
These results suggested that as brome continues to spread in the Great Plains, existing cordgrass patches will disappear with higher frequency, and new patches will appear with less frequency than in prairies free of brome. Although these data are compelling, they do not constitute proof that brome is directly responsible for the disappearance of cordgrass. However, Forrest completed a series of experiments to examine the competitive interactions between brome and cordgrass (see below).
Based on our experimental research on the movement of a specialist cordgrass planthopper (Prokelisia crocea) and its primary parasitoid (Anagrus columbi), we anticipate drastic consequences associated with the increase in prevalence of brome in the matrix and the reductions in cordgrass patch size and number. Relative to native matrix habitats (e.g., mudflat or native grasses), brome makes small cordgrass patches function like sieves for the two insect species. Planthoppers and parasitoids leak out of these patches at such high rates that extinction quickly follows (Cronin & Haynes 2004, Cronin 2007).
Mechanisms Responsible for the Spread of Brome:

Following the GIS survey, Forrest, along with the assistance of one of our NSF-REU students, Brian Thompkins (see photo), conducted a field experiment in North Dakota to determine the mechanisms responsible for smooth brome invasion. The experiment involved reciprocal transplants of brome and cordgrass seeds and seedlings into different tall-grass prairie habitat types (cordgrass, brome and a mixture of native grasses). In addition, the two plant species were either planted separately or together, and with and without thatch. The objectives of the experiment were to (1) test whether brome is capable of tolerating the abiotic conditions associated with each habitat type (i.e., does the abiotic environment limite the spread of brome?) and (2) test whether brome is a superior competitor to cordgrass.
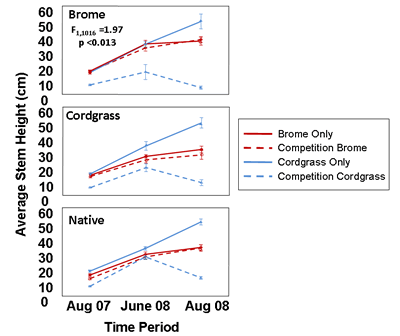
Forrest's results indicated that germination of both species varied with habitat type, but averaged across habitats, cordgrass seed germination was 1.5 times higher than smooth brome germination. However, by the end of the two year experiment, brome had an average of approximately 20% more stems than cordgrass regardless of whether the two plant species were grown along or together. In addition, when grown alone, cordgrass grew taller than brome, but when grown together, cordgrass stem heights decreased by approximately 70% regardless of habitat (Fig. 3). Finally Forrest found that during the 2 years in which this experiment was conducted, transplanted cordgrass seeds or seedlings failed to flower but flowering of brome occurred across all habitat types and in the presence or absence of cordgrass. Overall, these data suggest that once brome becomes established, it is a better competitor than cordgrass and can potentially displace cordgrass patches. Results from this study are described in more detail in Forrest's dissertation (Dillemuth 2012).
In a subsequent large-scale study, Forrest examined how the invasion of smooth brome has impacted the arthropod community associated with cordgrass. His work spanned multiple spatial scales, from individual patches, to prairie fragments, and to patchworks of prairie fragments (i.e., watersheds). Specifically, he examined the relationship between percent coverage of brome and its impact on arthropod richness, diversity, the relative abundance of specialist and generalist herbivores, and other aspects of community structure. As above, results from this study are described in more detail in Forrest's dissertation (Dillemuth 2012).





