Evolution and Ecology of Invasive Plant Species
Latitudinal Variation in the Interaction Between an Invasive Plant Species and Its Herbivores: Implications for Invasion Success
Project Overview

Biologists have recently recognized that contemporary evolution may play an important role in successful species invasions. For plant species, it is widely expected that latitudinal clines should evolve with regard to palatability and defense traits (palatability should increase and defenses decrease with increasing latitude). However, this ecological paradigm has never before been applied to species that are involved in continent-wide invasions. Moreover, no one has ever examined how interactions between invading plants and closely related native plants, via their shared herbivores, might vary across broad latitudinal gradients. For the past several years, Laura Meyerson (Natural Resources Science, University of Rhode Island), two past PhD students, Ganesh Bhattarai (Kansas State University), Warwick Allen (Lincoln University, New Zealand), and I have been addressing these issues with the cosmopolitan invasive grass, Phragmites australis (common reed; Fig. 1). This plant species is native and widespread in Europe and North America, but a European genotype has been spreading rapidly in North America over the past 100 years. This study system affords us the unique opportunity to examine parallel cline formation between native and invasive genotypes along the same latitudinal gradient, and assess how differences along this gradient could affect invasion success.
An informational (tri-fold) pamphlet on this research project is available as a PDF file in English, French, Spanish and Norwegian.

Background
P. australis represents a system for studying geographic and clinal variation in palatability, defense levels, and herbivore pressure; and how the invasion process is affected by this type of variation (Meyerson et al. 2016). Key features that make this an ideal system include, the co-occurrence of native and invasive genotypes across a wide (continental) range, a well documented history of invasion in North America, and a well known herbivore fauna associated with this plant in both native and introduced ranges. We provide a brief description of this system below.
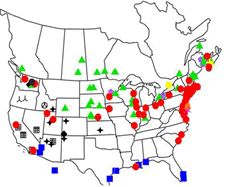
In the 1800s, botanical records described P. australis as being rare or not common, but by the early 1900s, it had become common and was spreading (Saltonstall 2002). However, in the ensuing years, P. australis spread throughout the U.S. and Canada and is now recognized as a dominant plant species in disturbed wetlands. The recent increase in P. australis, particularly along the east coast of the U.S., has been attributed to the introduction of an aggressive Eurasian genotype (Saltonstall 2002).
Based on chloroplast DNA analysis of fresh tissues and herbarium records, Saltonstall (2002) identified 27 distinct haplotypes worldwide, 11 of which were endemic to North America. These native haplotypes are found over much of North America, excluding the extreme southeast (see Fig. 2). Two additional haplotypes found in North America and elsewhere were designated haplotypes M and I. Haplotype M was the most common haplotype in present-day samples from North America, Europe and Asia. Haplotype M is considered invasive and first appeared in records from wetlands in the northeast, but has subsequently spread west (Saltonstall 2002, Lambertini et al. 2006, Meyerson et al.2010). Several additional native and invasive genotypes have since been discovered in North America (see Meyerson and Cronin 2013).
Project Objectives

The first phase of our research involved two main objectives. Our first objective was to test whether P. australis has undergone rapid evolution to form a cline with regard to traits associated with its nutritional condition and interactions with herbivores (palatability, defense levels). Our second objective focused on quantifying differences in palatability, defense levels, and herbivore pressure between sympatrically occurring native and invasive genotypes of P. australis, and evaluating how these differences may affect invader success. Using field surveys and common-garden experiments, the following specific questions were addressed. (1) Does invasive P. australis exhibit a latitudinal cline in nutritional condition, defense levels, and herbivore pressure that is parallel to the clines found for the same genotype in its native range (Europe) and with sympatrically occurring native genotypes (North America)? (2) Is invasion success associated with the invading genotype being less palatable, better defended, or having lower herbivore pressure than the sympatrically occurring native genotypes? (3) Is the geographical, latitudinal or within site (native vs. invasive genotype) variation in P. australis nutritional condition, palatability to herbivores, and defense levels genetically based? (4) Do native and invasive populations of P. australis interact through apparent competition; i.e., via shared herbivores?

Research Results
1) Biogeography of a plant invasion: plant-herbivore interactions.
Cronin, J. T., L. A. Meyerson, G. Bhattarai and W. Allen. 2015. Biogeography of a plant invasion: plant-herbivore interactions. Ecology 96: 1115-1127.
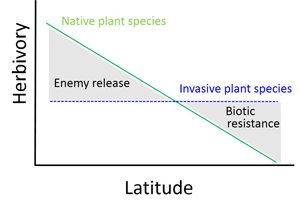
Theory predicts that native plant species should exhibit latitudinal gradients in the strength of their interactions with their herbivores. We hypothesized that if an invasive plant species exhibits a different latitudinal gradient in response to herbivores (e.g., a non-parallel gradient), it can create large-scale heterogeneities in community resistance/susceptibility to the invasive species (Fig. 4).
We conducted a study of latitudinal variation in the strength of herbivory and defenses of native genotypes of Phragmites australis in North America (NA) and Europe (EU) and European invasive genotypes in NA (Cronin et al. 2015). Within NA, we tested whether 1) invasive genotypes are better defended and suffer less herbivory than co-occurring native genotypes, 2) herbivory and defenses of native P. australis decreases with increasing latitude; and 3) invasive genotypes exhibit either no latitudinal gradient, or a non-parallel latitudinal gradient in herbivory and defenses compared to native genotypes. For the European genotypes, we tested two additional hypotheses: 4) defenses, nutritional condition and herbivory differed between the native (EU) and invasive ranges (NA) and 5) latitudinal gradients in defenses and herbivory were similar between ranges.
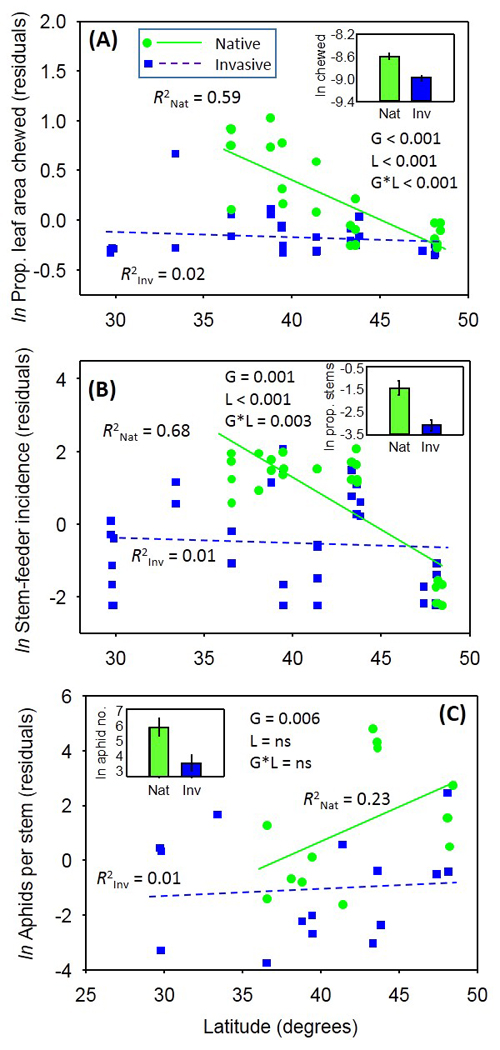
Within NA, chewing damage, internal-feeding incidence and aphid abundance were 650%, 300% and 70% lower, respectively, on invasive than native P. australis genotypes. Genotypes in NA also differed in nutritional condition (%N, CN ratio), but there was little support for invasive genotypes being better defended than native genotypes. For the European genotypes, herbivory was significantly lower in the invaded than native range, supporting the enemy-release hypothesis. Defense levels (leaf toughness, total phenolics) and tissue %C and %N were higher in the invaded than native range for European genotypes. Because all herbivores guilds exhibited a greater preference and caused more damage to native than invasive genotypes, we conclude that efforts to use insect herbivores for invasive P. australis management is likely to be detrimental to the relatively rare native genotypes of P. australis (see also Cronin et al. 2016, Bhattarai et al. 2016).
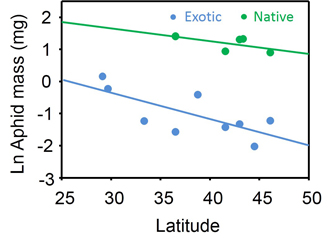
Overall, latitudinal gradients in P. australis nutritional condition, defenses and herbivory were common. Interestingly, chewing damage and stem-feeder incidence decreased with latitude for native P. australis genotypes in NA and EU, but no latitudinal gradients in response to herbivores were evident for invasive genotypes in NA (Fig. 5). Non-parallel latitudinal gradients in herbivory between invasive and native P. australis suggest that the community may be more susceptible to invasion at lower than at higher latitudes. Our study points to the need for invasion biology to include a biogeographic perspective.
2) Common-garden experiments to determine if latitudinal patterns are genetically based.
Bhattarai, G. P., L. A. Meyerson J. Anderson*, D. Cummings*, W. J. Allen and J. T. Cronin. 2017. The biogeography of a plant invasion: Genetic variation and phenotypic plasticity in latitudinal clines for plant-herbivore interaction traits. Ecological Monographs.87: 57-75
Bhattarai, G. P. 2015. Biogeographical approaches for studying species invasion. PhD Dissertation. Louisiana State University, Baton Rouge, Louisiana, USA.
The above survey is informative regarding the large-scale variation in P. australis-herbivore interactions, but controlled experiments were required to ascertain whether the observed variation is attributable to genetic differences among sites, or a consequence of phenotypic plasticity in response to an environmental gradient. At both LSU and URI, we established replicate common gardens that were populated with P. australis genotypes from sites across a 17° latitudinal range in North America (Fig. 6). We tested whether native and invasive lineages exhibited different genetically based latitudinal clines in defenses, nutritional condition and palatability to their herbivores (the aphid Hyalopterus pruni and the fall armyworm Spodoptera frugiperda). We also tested whether invasive genotypes are more phenotypically plastic than natives and whether plasticity varies with latitude.

Although invasive genotypes did not exhibit higher defense levels (leaf toughness, phenolics, % carbon), they were considerably less palatable to H. pruni and S. frugiperda than native genotypes (Fig. 7).
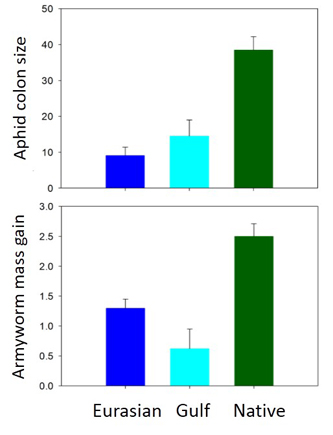
Genetic-based latitudinal clines were evident for both native and invasive lineages and for all defenses, nutrients and at least one palatability trait for each herbivore (e.g., leaf toughness, water content, and H. pruni colony growth decreased with increasing latitude for both lineages; Fig. 8).
In 36% of the cases where clines were evident, they were non-parallel between the two lineages. These data suggest that clines in the invasive genotypes of P. australis evolved within the past ~100 years. Moreover, our study showed that the occurrence and direction of latitudinal clines in plant traits were commonly dependent on where the study was conducted (north or south), indicating strong phenotypic plasticity in these genetic-based clines.
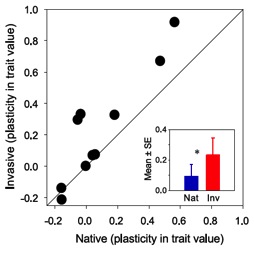
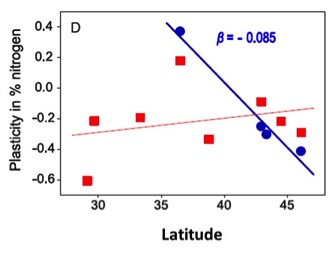
Finally, traits for invasive genotypes of P. australis were 2.5 times more plastic than traits for native genotypes (Fig. 9). Interestingly, plasticity for native but not invasive genotypes was strongly dependent on latitude of origin (Fig. 10). Such spatial heterogeneity within and between lineages with respect to their interactions with herbivores has the potential to generate substantial spatial variability in biotic resistance that can have important implications for the establishment and spread of invasive genotypes and species.
3) Apparent competition between native and invasive genotypes of P. australis.
Apparent competition, an indirect species interaction via shared natural enemies, has been shown to play an important role in shaping the structure and dynamics of natural communities. However, its importance in driving species invasion has not been fully explored. Moreover, latitudinal gradients in species diversity and biotic interactions can be expected to create similar spatial variation in the strength of apparent competition between interacting species. Ganesh Bhattarai and I performed replicated field experiments at four sites along the Atlantic Coast of the US to assess the presence and strength of apparent competition between sympatric native and invasive genotypes of P. australis. We also tested the hypothesis that the strength of this interaction declines with increasing latitude.
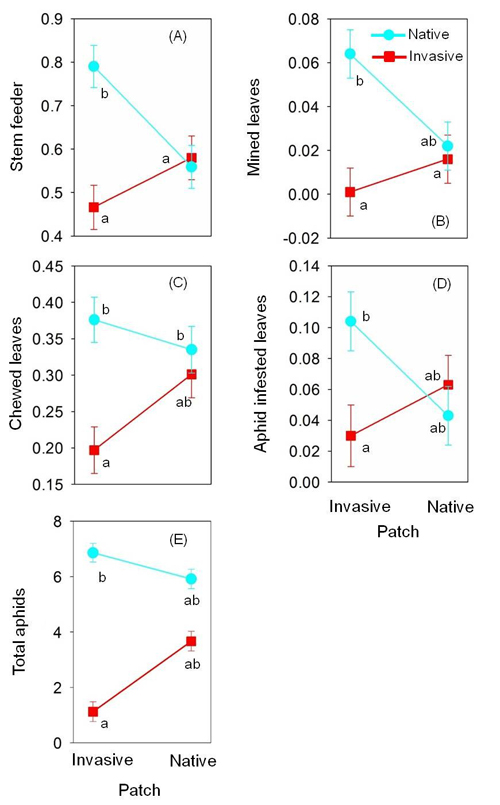
Within each site, native and invasive plants of P. australis were cross-transplanted between co-occurring native and invasive patches in the same marsh habitat and herbivore damage was evaluated by the end of the growing season. For native plants, the incidence of internal-feeding and leaf-mining herbivores was 43% and 191% higher, respectively, when transplanted into invasive than native patches (Fig. 11). Chewing damage and aphid infestation levels were equivalent on native plants in each patch type. These data suggest that invasive P. australis has a negative effect on native P. australis via increased internal-feeder incidence, indicative of apparent competition. In comparison, transplanted invasive plants did not experience a significant change in herbivory between native and invasive patches for any herbivore guild. Consequently, apparent competition appears to be asymmetric.
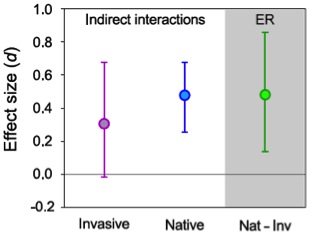
The strength of apparent competition between native and invasive P. australis, as measured by the mean effect size (Hedge’s d), is presented in Fig. 12. The 95% CIs for the mean effect of invasive genotypes on native genotypes did not overlap zero but they did overlap with zero for the reciprocal interaction. Overall, the strength of the negative effects of invasive genotypes on native genotypes was 57% higher than the strength of the negative effects of the native genotypes on the invasive genotypes. Finally, the strength of apparent competition of the invasive genotypes on the native genotypes was statistically indistinguishable from the strength of enemy release for the invasive genotype (Fig. 12).
As predicted by theory, but never before tested, the intensity of apparent competition declined with increasing latitude. These results suggest that apparent competition mediated by internal-feeders (primarily, galling flies) might be one of the mechanisms enhancing the invasion of introduced genotype of P. australis in Atlantic Coast of the United States. Invasion success may also vary with latitude, being more likely at southern latitudes where apparent competition is stronger. More details regarding this project can be found in Bhattarai (2015) and Bhattarai et al. (2017).
References
- Howard, R. J., S. E. Travis, and B. A. Sikes. 2008. Rapid growth of a Eurasian haplotype of Phragmites australis in a restored brackish marsh in Louisiana, USA. Biological Invasions 10:369-379.
- Lambertini, C., M. H. G. Gustafsson, J. Frydenberg, J. Lissner, M. Speranza, and H. Brix. 2006. A phylogeographic study of the cosmopolitan genus Phragmites (Poaceae) based on AFLPs. Plant Systematics and Evolution 258:161-182.
- Meyerson, L.A., A. Lambert, K. Saltonstall. 2010. A tale of three lineages: expansion of common reed (Phragmites australis) in the U.S. southwest and Gulf Coast. Invasive Plant Science and Management 3:515-520.
- Pellegrin, D. and D. P. Hauber. 1999. Isozyme variation among populations of the clonal species, Phragmites australis (Cav.) Trin. ex Steudel. Aquatic Botany 63:241-259.
- Saltonstall, K. 2002. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proceedings of the National Academy of Sciences of the United States of America 99:2445-2449.
- Tewksbury, L., R. Casagrande, B. Blossey, P. Hafliger, and M. Schwarzlander. 2002. Potential for biological control of Phragmites australis in North America. Biological Control 23:191-212.





