Host-Parasitoid Spatial/Landscape Ecology
Matrix Composition Affects Dispersal and Population Dynamics
Our extensive research has shown that the matrix does matter for P. crocea and its parasitoid, A. columbi (Cronin 2003b, 2004, Haynes & Cronin 2003, Cronin & Haynes 2004, Haynes & Cronin 2006, Cronin 2007, Haynes et al. 2007, Cronin 2007, Reeve et al. 2008, Reeve and Cronin 2010, Cronin and Reeve 2014). We found that planthopper emigration rates were significantly higher for cordgrass patches embedded in a matrix composed of brome than mudflat, and the parasitoids showed a similar response. For both species, the brome edge was “soft” (allowing free movement into the matrix) and the mudflat edge was “hard” (functioning as a barrier to emigration). Interpatch movement behavior also was strongly influenced by matrix composition.

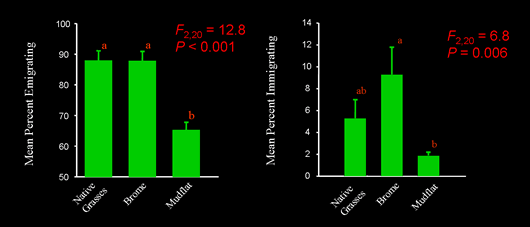

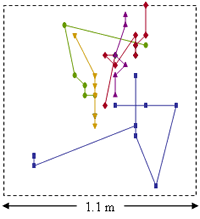
In a brome matrix, planthoppers exhibited tortuous movement pathways and short step lengths, whereas in a mudflat matrix, pathways were linear and step lengths were long. Planthopper movement in both matrix types was well fit by a diffusion model that allowed for heterogeneous dispersal rates (see Cronin et al. 2000). However, overall rates of diffusion and the proportion of “fast dispersers” were considerably higher for the mudflat than the brome matrix. Less information is available on A. columbi interpatch movement. Overall, the planthopper and parasitoid exhibit very similar redistribution patterns in the 2 matrix types. In nature where host-plant patches are spatially aggregated, tortuous movement pathways, like those exhibited by planthoppers in brome, are expected to result in greater immigration rates (Haynes 2004, Haynes & Cronin 2006). In fact, we found that emigrating planthoppers and parasitoids were 5-6 times more likely to colonize cordgrass patches when those patches were embedded in brome as compared to mudflat (Cronin & Haynes 2004). Thus, brome functions like a low resistance matrix (one that facilitates dispersal) for the planthopper and its parasitoid and mudflat functions like a high resistance matrix (one that impedes dispersal).
Matrix composition also had a significant impact on the spatial and temporal dynamics of the host and parasitoid. Our 7-generation survey revealed that occupancy rates and densities of both species were higher in patches surrounded by mudflat than brome. For the planthopper, the risk of local extinction increased with the proportion of the surrounding matrix (within 3 m) that was brome. These patterns were directly attributable to the higher rates of emigration loss (and edge permeability) for patches bordered by brome than mudflat.


We also performed a landscape-level field experiment in which replicate cordgrass networks were embedded in a matrix composed of either mudflat or smooth brome. The rate of colonization of patches separated by 50-60 m is on the order of 0.1-0.01% per generation (Cronin & Haynes 2004), indicating that these experimental landscapes (3 replicates per matrix type) are large from the perspective of the host and parasitoid. Mark-recapture experiments with these 2 species revealed that the rate of movement among patches was 3-11 times higher in brome than in mudflat. Over the course of 5 generations, planthopper and parasitoid densities/patch were on average ~50% lower and spatially 50-87% more variable for patches embedded in a brome as compared to a mudflat matrix. A brome-dominated landscape also promoted extinction rates per patch that were 4-5 times higher than the rates per patch in mudflat habitat: the effect was more acute for the parasitoid. In fact, during generation 5 of this study (Aug., 2004), regional extinctions (i.e., extinctions in all 15 patches of a network) took place in all 3 brome networks (Cronin 2007). However, the 3 mudflat networks remained viable (patch vacancies < 30%).
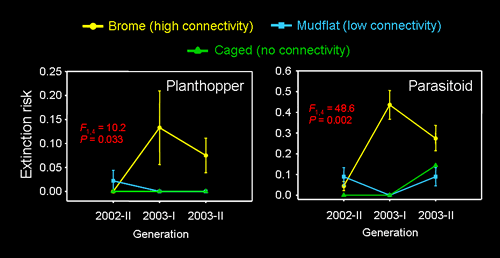
We suggest that the differences in population dynamics between networks of patches in brome and those in mudflat were driven by underlying differences in interpatch dispersal. This was the first experimental study to reveal that matrix composition, in particular, the presence of an invasive plant species affects the spatial and temporal dynamics of an herbivore and its natural enemy.





